Goldfish as a Model for Understanding Learning and Memory: More Complex Than You Think
Part of the Young Naturalist Awards Curriculum Collection.
Purpose
The United States and the European Union are pouring hundreds of millions of dollars into research on understanding how the human brain functions. As several key aspects of learning and memory are shared by all animals (Kandel, 2006), researchers are studying different kinds of animals, from simple organisms to humans, to help explain learning and memory in the human brain. Most research has occurred in mammals, but some attention has been placed on fish as an animal model to help delineate the types of memory needed to complete spatial tasks such as navigation. Spatial cognition, a form of explicit memory that involves navigating around spaces, is found in animals with higher-level brain function, specifically those animals with an area of the brain called the hippocampus, the part of the cortex that stores explicit memory (Kandel, 2006). Experiments on rats led scientists to conclude that rats, like humans, can create in their brains a “comprehensive map” of the room in which experiments are performed, called a cognitive map, in order to navigate to a goal (Tolman, 1946). Unlike rats and humans, the fish brain does not have a cortex and consequently no hippocampus in which to store explicit memory, leading scientists to conclude that fish cannot create a cognitive map. However, scientists have found that fish do use spatial cognition, relying on allocentric (extra-maze) cues to navigate, and that there are areas of the fish brain that are analogous to the hippocampus (Salas et al., 2006). These findings suggest that fish have a higher level of functioning than is commonly believed (Brown, 2015) and perhaps have the ability to form a cognitive map. Moreover, as the first group on the vertebrate branch from which mammals descended, this finding would indicate that complex brain functions began to develop hundreds of millions of years ago.
This science project, which is in its third year, attempts to add to the body of research on fish learning and memory in a unique way. First, the length of long-term memory in fish in general has not been delineated, and certainly not in goldfish, though there have been anecdotal reports of long-term memory in certain species of fish, such as salmon returning home to spawn. Second, although the use of allocentric (extra-maze) and egocentric (intra-maze) cues by goldfish after training is well documented(Salas et al., 1996), delineating the time frame during training when goldfish switch from allocentric to egocentric cues to learn and memorize a task, has not been done. This time frame would serve to indicate when learning is converted from short-term memory to long-term memory. Third, this experiment uses different and more difficult mazes than the T-mazes, plus mazes, or small square box mazes used by researchers. The design of the mazes gives the goldfish several options in deciding where to navigate, and allows for the observation of behaviors relevant to determining whether the fish use place strategy (spatial cognition) or response strategy (rote memory) during the learning process, to an extent not previously documented or researched.
The exciting impact of research on goldfish would be to rewrite the biology textbooks about the place of fish in evolution as the starting point for the development of a higher-functioning brain, in particular the use of spatial cognition. Moreover, the ultimate goal would be to use these findings to understand learning, memory, and memory loss in humans, with the hope of one day unlocking the mystery of Alzheimer’s disease, a disease of spatial cognition.


Research Questions
- How long is the long-term memory of goldfish?
- How do the goldfish learn a spatial task? Specifically, which navigational strategies—place strategy (spatial cognition) or response strategy (rote memory)—did the goldfish use to learn to find the food in the maze during training?
Background
I have owned goldfish and other types of fish for many years and am always astonished that they seem to recognize me when I come to feed them. My research into goldfish learning and memory started with my first science fair experiment three years ago to determine whether goldfish could learn to find food in a maze. In my literature review, I was disappointed to find that goldfish memory was frequently and often deridingly characterized as lasting three seconds (Osborne, 2010). However, my initial research showed that not only did goldfish learn quickly, but their memory was certainly longer than three seconds. My second-year experiment confirmed that goldfish can learn quickly, and showed that goldfish can recall memories for at least one month. Several months passed while I thought about what my next experiment, during which time I retested the goldfish on their memory of the maze from my second-year experiment. I was impressed that the goldfish located the food in the maze faster than on their last day of training. As a result, I became intrigued with two questions: (1) What is the duration of goldfish memory? and (2) How are goldfish able to recall the location of food in the maze after not being in the maze for several months?
A significant amount of research has been done on goldfish learning and memory. Researchers in Spain have found that certain areas of the goldfish brain, called the medial and lateral telencephalon, correlate to regions of the human brain that deal with memory—the amygdala and hippocampus, respectively—by ablating parts of the goldfish brain and assessing its recall of a learned behavior in a plus maze (Salas et al., 1996). They found that spatial cognition in goldfish was dependent on a functioning lateral telencephalon, but response strategy navigation could occur without a functioning telencephalon (Rodríguez et al., 1994; López et al., 2000; Salas et al., 2006). They concluded that dual, parallel learning systems exist in goldfish: namely, a higher-level spatial cognitive strategy using allocentric cues and a lower-level response strategy (López et al., 2000; Vargas, 2004). The purpose of their studies was to delineate the functions of the goldfish brain and to provide evidence for the evolution of spatial cognition among various animal species, starting with fish (Rodríguez et al., 2002).
Further review of the scientific literature did not reveal any studies addressing the duration of long-term goldfish memory, the use of a complicated maze, or what learning strategies goldfish use during the training process. As the type of memory experiments proposed here can take more than one year to complete, it is not surprising that studies characterizing goldfish memory have not been done. In addition, the uniqueness of evaluating how goldfish learn during training was not addressed in the literature from Spain, which focused on testing the goldfish after training was complete. For example, the previous research considered how well the goldfish performed on reversal testing, in which the goal was changed after training, and not on which navigational strategy they used during training. Their research did not evaluate if and when the goldfish switched from using allocentric to egocentric cues during training, or if goldfish were predisposed to using one strategy over another. Many of the experiments done on goldfish in the past used classical conditioning and dealt with implicit memory, which does not require conscious attention for recall because the behaviors are usually reflexes (Kandel, 2006). Not wanting to shock the fish in these experiments, I did not use classical conditioning; instead, I used positive reinforcement with food.
In thinking about how goldfish learn the route to the food in the maze during training, I hypothesized that, in the short term, the goldfish would use landmarks in the room, or allocentric cues, to remember the location of the food, using place strategy (spatial cognition) to navigate to the goal. As the goldfish would be trained daily for 28 days, I suspected that consistently turning left into the food cubicle would become routine, predictable, and rote in the long term, leading the goldfish to rely on response strategy (rote memory) to navigate to the goal. The use of allocentric cues during the learning process of a spatial task would indicate that goldfish do have a complex component to their brains, and that spatial cognition is not limited to higher mammals and humans simply because they have a cortex. Therefore, using goldfish (Carassius auratus) as an animal model to understand learning and memory is relevant and important to delineating the evolution of spatial cognition as well as to potentially understanding diseases of memory.
Hypotheses
If goldfish have long-term memories lasting up to six months, then goldfish trained to find food in a maze will, after intervals as long as six months, be able to recall the location of the food in the maze significantly faster than on the first day of training. During training, goldfish will initially learn to find food in a maze by using place strategy (spatial cognition), noted by a greater number of right turns than left turns in the test mazes. By the end of the training period, goldfish will change to a response strategy (rote memory), noted by a greater number of left turns than right turns in the test mazes. Goldfish in the short term will rely on place strategy (spatial cognition) and allocentric cues (extra-maze cues) to solve the task, but will shift to a response strategy (rote memory) once the task becomes repetitive and predictable.
Research Methods
Materials
- Forty large goldfish (Carassius auratus) of various breeds
- Omega 1 small fish food pellets
- Four clear plastic storage boxes (73.6 cm x 45.7 cm x 15.2 cm; 38.8 liters) for mazes
- Several large aluminum baking pans, used as partitions in the maze
- Five-foot-tall cardboard box for a surround
- Flat white and dark brown spray paint
Mazes
I made four different types of mazes, cutting the corners off the aluminum pans to make partitions for the cubicles and attaching them to the clear plastic storage boxes with a hot glue gun.
Clear Training Maze

The first type of maze was the clear maze used for training goldfish in last year’s experiment (Figure 1). The clear maze was used to determine the duration of long-term memories in goldfish in Experiment 1 and to train the new goldfish for Experiment 2, called the “clear maze group.”

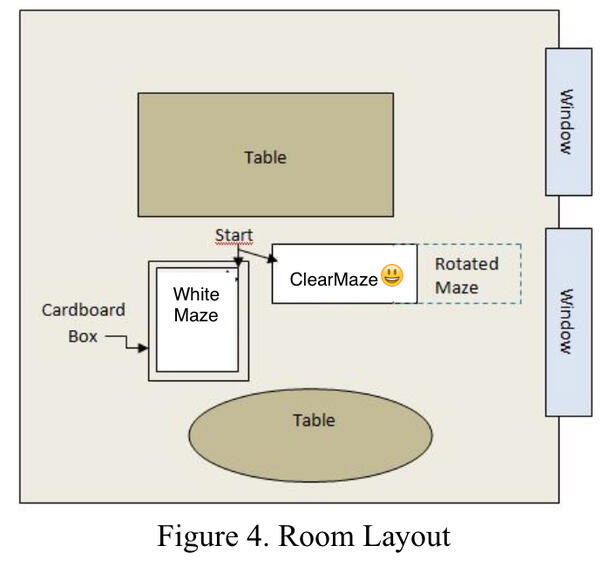
The goldfish trained in the clear maze were exposed to allocentric (extra-maze) cues in the room used to conduct the experiment (Figure 4, room layout). One salient allocentric cue was the five-foot-tall cardboard box, with the right half painted dark brown and the left half painted flat white (Figure 5), that faced the clear training maze and two of the three test mazes (Figures 3b, 3c).
White Training Maze (Used by the Control Group)
The maze for the control group (Figure 2) had the same design as that used to train goldfish in Experiments 1 and 2, except that it was painted flat white and was surrounded by a five-foot-tall cardboard box, the interior painted flat white, to remove almost all of the allocentric cues used by the goldfish to navigate except those subtle cues related to imperfections in the aluminum partitions and the cardboard box, as made visible by the overhead light.
Test Mazes
Three test mazes (Figures 3a, 3b, 3c) were used intermittently during the training period. The first test maze (Figure 3a) was the clear training maze, but surrounded on all four sides by the five-foot-tall cardboard box painted white on the inside. The purpose of the box was to remove allocentric cues and observe whether the goldfish relied on egocentric cues to navigate.
The second test maze (Figure 3b) was a double-chamber maze with two right cubicles and two left cubicles, designed to change the egocentric cues while the allocentric cues remained unchanged. The purpose was to observe whether the change in egocentric cues fooled or confused the goldfish into entering the cubicles closest to the start position, and whether the goldfish relied on allocentric cues to navigate.

The third test maze (Figure 3c) was an opposite-start shortcut maze, designed to change the egocentric cues while the allocentric cues remained the same. The purpose was to disorient the fish by placing them in a different start position in the maze and to observe whether the goldfish relied on allocentric cues to navigate. The shortcuts were openings on either side of the end cubicles that allowed the goldfish to enter them directly without having to turn. The purpose here was to observe whether the goldfish could recognize a second entrance to the cubicle they believed the food was in.

Procedure
Experiment 1: Goldfish Long-Term Memory Span
- The trained fish from last year’s experiment were placed in the start cubicle of the clear training maze using a small net (Figure 1). Food pellets were placed in the left cubicle of the clear training maze. The experimenter quietly backed away from the maze to record the times and responses.
- The time in seconds from the moment the fish exited the net into the start cubicle to the moment the fish ate the food in the left cubicle was recorded. The goldfish were tested at two-, three-, four-, and six-month intervals to determine whether they recalled the location of the food after not performing the task for two, three, four, and six months.

Experiment 2: Goldfish Learning (Training) and Comparison
- Sample Size:Forty goldfish of different breeds were randomly assigned to either the training/testing group, called the clear maze group (n = 30), or the control group (n = 10).
- Clear Training Maze (n = 30): The goldfish were placed individually into the start cubicle of the clear maze once a day for 28 consecutive days (Figure 1). The time it took them to find the food in the left cubicle was recorded, as well as any other observations, including the route taken (left or right) and which cubicle (left or right) the fish initially entered.
- White Maze for the Control Group (n = 10): The goldfish were placed individually into the start cubicle of the white maze once a day for 28 consecutive days (Figure 2). The time it took them to find the food in the left cubicle was recorded, as well as the route taken and the initial cubicle entered. The goldfish in this group served as the control because they had to use response strategy (rote memory) for navigation, as there were few, if any, allocentric cues present beyond the texture of the painted aluminum partitions and the imperfections on the inside of the cardboard box surround.
Experiment 3: Testing During Training to Determine Place vs. Response Strategy in Learning
- Thirty goldfish were trained in a clear maze. In the clear training maze, a decal (a smiley face) marked the exact location of the food and was visible through the clear floor of the maze (Figure 1). The test mazes (Figures 3a, 3b, 3c) were then rotated 180° so that the cubicle that earlier in the day had held the food (Figure A) was now in the same location as the empty cubicle (Figures B, C). The goldfish were randomly assigned to one of the three test mazes. The mazes were rotated 180º to test whether the fish were using a response strategy. No food was placed in any of the test mazes to avoid giving olfactory clues to the fish to turn in a certain direction. The time the fish took to reach the goal was recorded, as well the route taken and which cubicle was initially entered.
- On Days 3, 7, 11, 15, 19, 23, and 27, the fish who had gotten food earlier in the day in the training maze were placed and observed in the test mazes, for seven trials per fish. These intervals were chosen based on last year’s experiment, which showed that goldfish learned to find the food in the maze by the third day, and made fewer mistakes and had faster times on the days that followed.
Variables
Dependent Variables:
- Time in seconds to find the food in the training maze (Experiments 1, 2)
- Correct food cubicle (left cubicle in training maze) (Experiment 2)
- Whether fish turned to the right cubicle (place strategy) or the left cubicle (response strategy) in the three testing mazes (Experiment 3)
Independent Variables:
- Time in months absent from maze (Experiment 1)
- Days of training (Experiments 1, 2)
- Three different testing mazes with 180º rotation (Experiment 3)
Results
Experiment 1: Goldfish Long-Term Memory Span
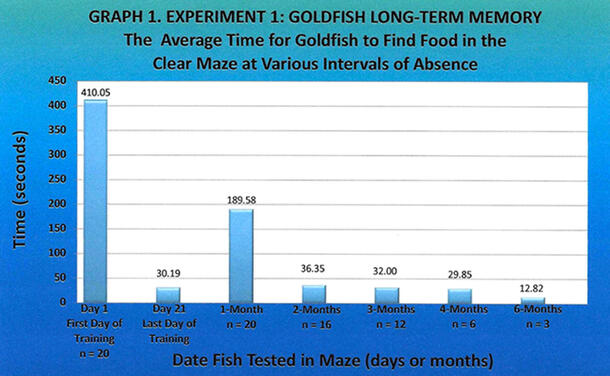
The goldfish used in last year’s experiment were retested at two-, three-, four-, and six-month intervals in the original clear maze (Figure 1). Their average times were quick and improved over time: 36.35, 32.00, 29.85, and 12.82 seconds, respectively, at two, three, four, and six months. The average time at one month was the longest, at 189.58 seconds (Graph 1). All five average times were significantly improved compared to the first day of training (410.05 seconds), at p < 0.0005 using a one-sided lower-tail t-statistic.
Experiment 2: Goldfish Learning (Training) and Comparison
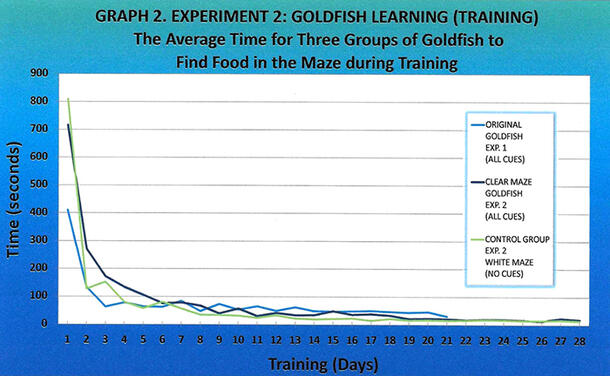
For all of the fish tested—the goldfish trained for last year’s experiment, the goldfish trained in the clear maze, and the control group—the average time it took the goldfish to find the food in the maze was very similar, showing rapid improvement by the second day of training (Graph 2). By the ninth day, each group had an average time of less than 60 seconds. Overall, the control group was quickest at finding the food in the maze.
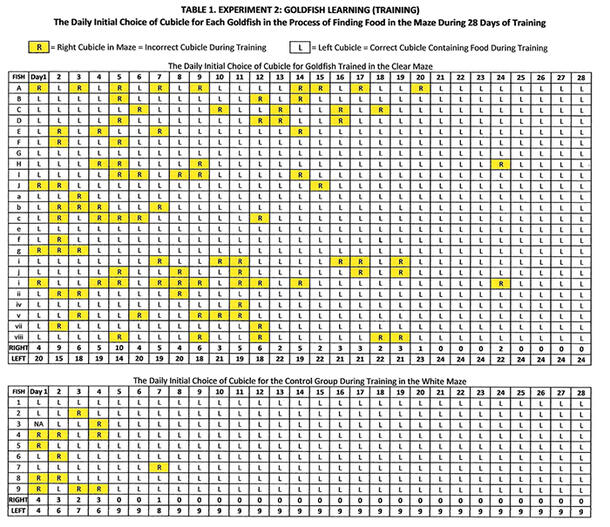
The initial cubicle entered by all the fish during training was recorded daily for 28 days (Table 1). In the training maze and the white maze, the food was consistently located in the left cubicle (Figures 1, 2). The fish in the control group were correct 94% of the time in the white maze, compared to the fish trained in the clear maze, who were correct 86% of the time in the white maze (Table 1). The control group made fewer mistakes after Day 7, compared to Day 20 for the fish trained in the clear maze. The difference was statistically significant at p < 0.001 (Mann-Whitney U-Test, p = 0.00019, U-value = 135, Z-score = 3.5572).
Experiment 3: Testing During Training to Determine Place vs. Response Strategy in Learning
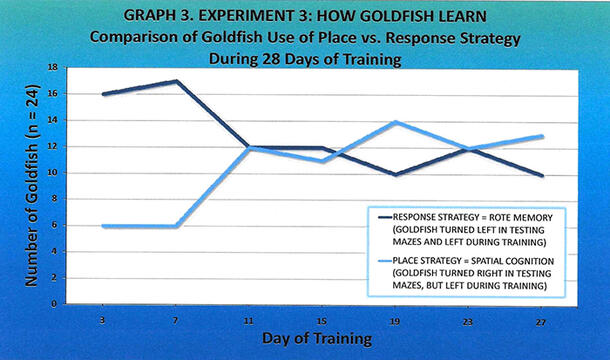
The initial cubicle entered by the fish in the rotated test mazes (Figures 3a, 3b, 3c) was recorded as place (the fish turned right, indicating a place strategy) or response (the fish turned left, indicating a response strategy) for each of the seven trials during the 28-day training period (Table 2). The number of response strategies declined (to 10 on Day 27, from 16 on Day 3) and the number of place strategies increased (to 13 on Day 27, from 6 on Day 3) during training (Graph 3). The change from response to place strategy was statistically significant at p < 0.05 (Mann-Whitney U-Test, p-value = 0.04746, U-value = 179, Z-score = 1.6689).
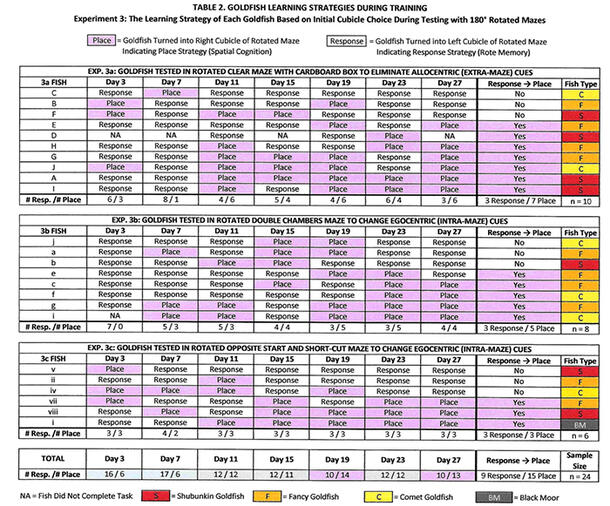
Although all the goldfish were from the same species, Carassius auratus, different breeds were used to help distinguish the fish during the experiment. Seven out of ten (70%) of the fancy goldfish used place strategy by the end of the 28-day training, in comparison to four out of seven (57% ) of the shubunkin goldfish, and 3 out of 6 (50%) of the comet goldfish (Table 2).
Discussion
Experiment 1: Goldfish Long-Term Memory Span
My hypothesis regarding goldfish having a long-term memory of at least six months was correct. The average time for the goldfish to find food in the maze after an absence of six months was 12.82 seconds, which was less than half of the time taken on the last day of training (30.19 seconds) 16 months earlier (Graph 1). These results suggest that the ability of goldfish to retain and recall an explicit memory is impressive considering its small brain size, short life span, and lowest place on the vertebrate hierarchy.
Of note, the one-month average time of 189.58 seconds was the highest of the five readings. There can be two explanations. First, the one-month time was an aberrancy. Although this is possible, I would use this explanation if, for example, the third-month time had been the longest, with the other four times being similar. The second explanation is that the fish began to lose their memory of finding the food in the maze because this was their first major length of time away from performing the task in the maze. As they no longer needed to find food in the maze because they were being fed in the fish tank, the relevance or importance of remembering the task was diminished. Simply, this was a memory that they did not need.
But ultimately, something happened in the maze for the goldfish to remember the location of the food. From my observations, when the fish returned to the maze after an absence of one month, they stayed near the start position, eventually moving slowly along the maze wall to the middle of the maze, when some cue spurred their memory, causing them to suddenly swim quickly to the left cubicle containing the food (Figure 1). As the maze at the middle point is symmetrical and the food was not visible, I wonder whether the fish noticed an allocentric (extra-maze) cue that triggered their memory of food being in the maze. This would be consistent with the findings discussed below, that fish rely on spatial memory to solve a task. In addition, as the number of months since performing the task increased, goldfish performance improved, suggesting that spatial memory becomes ingrained in the brain, perhaps with further protein synthesis that makes recall easier.
In summary, this data suggests that goldfish have a spatial memory of at least six months, and reinforcement of the task at the one-month period appears to be critical to establishing long-term memory.
Experiment 2: Goldfish Learning (Training) and Comparison
All three groups of goldfish—the trained fish from last year, the clear maze group, and the control group—were able to learn to find the food in the maze in comparable times (Graph 2). The fish were initially cautious when placed into the maze, especially on the first day, quietly observing the environment by looking up, swimming slowly along the right wall out of the start cubicle to explore the rest of the maze, and, finally, finding the food by trial and error, taking on average 12.35 minutes. By the second day, the fish cut their time by more than 60% for the clear maze group and more than 80% for the control group (Graph 2). By the end of one week, the average time for all of the goldfish to find the food in the maze dropped to 1.2 minutes, and by the end of four weeks, the average time was 17.21 seconds. These findings demonstrate that goldfish learn quickly and can improve their ability to complete a task when it is repetitive and reinforced positively with food.
In general, the goldfish frequently followed the right wall of the maze out of the start cubicle (the opening of the start cubicle faced the right wall) to explore their surroundings in the training maze (Figure 1). Once accustomed to the maze, most goldfish swam directly to the goal, taking the left or right route. Sixty-four percent of the clear maze fish preferred to take the right route, in comparison to 71% of the control group fish; this difference was not statistically significant. I observed that some fish, if they did not initially swim to the food cubicle because they were distracted by something, such as their reflections in the clear maze walls, or if they were swimming back and forth along the maze walls, would eventually circle back to the start cubicle, which was a familiar spot, and appear to “reset” their brains as to why they were in the maze. Then the fish suddenly headed directly to the food cubicle from the start position.
The control group had the fastest times for finding the food and the fewest mistakes in entering the correct food cubicle (left cubicle) during training, the latter dramatically noted by the absence of yellow highlight after Day 7 of training (Table 1). This finding is reflected in my observations that the control group was able to establish an association between food and the left cubicle seven to ten days before the clear maze fish established an association. Certainly, the control group faced none of the distractions encountered by the clear maze fish, such as fighting their reflections in the clear walls of the maze, or spending time observing the objects in the room (the allocentric cues). Although the control group had the fastest times and fewest mistakes, these results do not suggest that the control group fish were smarter than the clear maze fish, just more focused, without the distractions.
Experiment 3: Testing During Training to Determine Place vs. Response Strategy in Learning
My hypothesis that the goldfish would initially use place strategy in learning to find the food in the maze, then switch to response strategy as the task became routine, was wrong. The results showed the opposite (Graph 3). Seventy-three percent of the training/testing goldfish (the clear maze group) used response strategy initially, but by the end of four weeks of training, only 43% used response strategy. In contrast, only 27% of the training/testing goldfish used place strategy initially, but by the end of four weeks of training, 57% used place strategy. This shift from using response strategy (rote memory) to place strategy (spatial cognition) was statistically significant.
To explain these results, I believe that the goldfish initially used response strategy (turning left into both the training and the test mazes) because it was the easiest to remember (i.e., turn left in training, so turn left again in the rotated test maze later on). As the goldfish became more familiar with the location of objects in the room, including the brown and white cardboard box, a salient allocentric (extra-maze) cue, they began to use place strategy (turning left in the training maze and right in the testing maze) on Day 11 (Graph 3). In fact, I noticed that several goldfish would stop at the end of the test maze, look at the cardboard box, and then turn right. This switch from response strategy to place strategy makes sense because it would take several days, as the goldfish were in that part of the room for only a few minutes once a day, to observe and commit to long-term memory the placement of objects in the room. Moreover, the fish that used place strategy were able to pinpoint the location of where they thought the food would be (in the right cubicle), an area separated from the left cubicle by only ten inches. As a result, I believe that the ability of goldfish to learn quickly, observe and memorize their environment, and recall that information, is very impressive. Whether the goldfish would continue to use place strategy to solve the task after four weeks remains unknown; however, the trend is that they would.
In addition, my findings confirm research on the necessity of allocentric cues for goldfish spatial cognition and navigation. The five-foot-tall cardboard box used to surround the clear maze (Figure 3a) had a profound effect on the fish. In the beginning they seemed confused by the sudden lack of allocentric cues and studied their new environment intently, looking up at the ceiling, slowly swimming around the maze while peering through the clear walls, and spinning their upper bodies around while their tails remained stationary, all in an attempt to understand the altered setting. The goldfish tested in Figure 3a initially used a response strategy, indicating that, without allocentric cues to guide them, the fish relied on rote memory. By the end of four weeks of training, however, the goldfish used place strategy (spatial cognition) to navigate, making a strong case for a cognitive map in order for them to be able to navigate to the goal while, in a sense, blindfolded by the lack of allocentric cues.
In regard to the other two test mazes (Figures 3b, 3c), which changed egocentric cues, the effect of changing the non-salient cues was negligible. The double-chamber maze (Figure 3b) did not fool many fish into initially entering the cubicles closest to the start position. The fish knew that the relevant cubicles were at the far back of the testing maze. The opposite-start maze (Figure 3c) did not affect the ability of the fish to navigate to the back of the maze and did not appear to affect their decision to turn left or right. Many fish did use the shortcut into the response cubicle (left cubicle), but this may have been an artifact of the maze design (the opening of the start cubicle faced the left wall), as the fish followed the entire left wall out of the start position without stopping, swimming into the left cubicle without much thought.
Food was purposefully omitted from the test mazes (Figures 3a, 3b, 3c) to allow the goldfish to turn freely into the cubicle they thought contained the food. Two of the test mazes (Figures 3b, 3c) never contained any food pellets, and consequently the fish could not have been influenced by olfactory cues. The Figure 3a maze, the clear maze with the cardboard box surround, was the same maze used in training earlier in the day and did contain food pellets. Any uneaten food pellets were removed after each trial and before each testing session. I do not think that any lingering smell from the food remnants influenced the fish, because the number of goldfish turning into the right cubicle (which never had any food in it) increased and the number of fish turning into the left cubicle (which consistently contained food) declined over time during the 28-day training period.
The range of goldfish learning ability seemed to mirror what is seen with human learning. Most of the goldfish quickly learned to find the food in the maze. Some fish learned faster than others; some were easily distracted. One was very clever, namely Goldfish D: on Day 27, the last day of testing, Goldfish D swam to the end of the test maze (Figure 3a), but instead of entering the right cubicle, it only looked in, taking several minutes to determine whether any food pellets were present, and, when none were visible, it turned to face the left cubicle, looking in and again taking several minutes, but not entering. When nothing was visible, Goldfish D returned to the middle of the maze and sat there. This goldfish had previously been cautious, not entering any cubicle at the beginning of the seven testing trials, but did eventually enter the left cubicle on two subsequent days and then the right cubicle on another day, before deciding not to enter either cubicle on Day 27. This goldfish was the only one that exhibited this unique behavior, suggesting a high degree of sophistication in understanding that food was present in the open training maze but not necessarily in the closed test maze and, to avoid falling into a trap, acted cautiously while being tempted with the hope of finding food. On the other end of the spectrum, seven goldfish were not included in the statistical analysis because they were untrainable, did not seem to understand the task, and required more than two minutes to find the food by the end of training (the average time for the rest of the fish was less than 20 seconds). These fish either entered the cubicle without food or seldom entered the food cubicle, remaining near the start position or in another area of the maze.
During training, certain goldfish remained response-strategy users, while others predominantly used place strategy, suggesting a possible genetic component to strategy preferences. Most of the fish used both strategies during the seven testing trials, consistent with previous research indicating that goldfish have parallel navigation systems. However, individual fish appeared to have a preference over time as to which strategy to use (Table 2). Further analysis showed there were differences among the same species of goldfish: namely, 70% of fancy goldfish used place strategy by the end of four weeks of training, in comparison to 57% of shubunkin goldfish and 50% of comet goldfish (Table 2). I suspect that the data from the fancy goldfish had some impact on the conclusion, because the fancy goldfish accounted for 42% of the training/testing group and 70% of them used place strategy. Consequently, the results might have been stronger had I used only fancy goldfish in the experiment, but it does raise an interesting question about differences in learning ability and strategies among the same species of fish. This finding of differences within the same species of fish, aside from one study that noted differences within stickleback species (Odling-Smee & Braithwaite, 2003), has not been reported in goldfish. As a result, future experiments should be conducted with one breed of goldfish or with equal numbers of different breeds.
In summary, my hypothesis on goldfish learning was incorrect because I thought that goldfish would use rote memory to find the food in the maze over time as the task was repetitive. Instead, the goldfish used allocentric cues to navigate to the goal over time, giving themselves an opportunity to observe, memorize, and commit to long-term memory the location of objects in the room. I underestimated the ability of goldfish to keenly observe their surroundings, as they spend their time swimming in a glass tank, but goldfish are more complex animals than I thought. Even though these results were not expected, I am encouraged more than ever to study the learning capabilities of fish because, as the findings suggest, goldfish do rely on allocentric cues to navigate and appear to map their environment in their brains, making a strong case for fish having a cognitive map.
Conclusion
My research indicates that goldfish learning a spatial task switch from predominantly using a response strategy (rote memory) to predominantly using place strategy (spatial cognition). This switch starts at approximately ten days, after the goldfish have had time to observe and memorize the allocentric cues in the environment integral to developing a spatial map in their brains, known as a cognitive map. For the goldfish to turn in the opposite direction from where they had turned earlier in the day, during training, implies that goldfish are able to recall and use this cognitive map to swim to the goal. The ability of goldfish to recall their surroundings in this way indicates that goldfish have a higher or more complex component to their brains, perhaps something akin to place or grid cells in mammals. Moreover, goldfish are able to retain a spatial memory for at least six months, solving the task with increasing speed despite not performing the task for long periods of time. These findings are very exciting and compelling for further research on the spatial cognition of goldfish, which are more complex animals than previously believed.
Further Research
Spurred by my encouraging results—that goldfish develop a spatial memory while learning to solve a task and have long-term recall of that task—I would like to continue to test the spatial cognition of goldfish by constructing a larger and more geometrically complicated maze while altering the allocentric and egocentric cues. I would also continue testing the duration of long-term memories in goldfish. This research could be important because significant differences may occur in the goldfish brain between when it has a memory of a specific task and when it loses that memory.
Further research on fish would include looking for the equivalent of the place and grid cells found in mammals that are responsible for learning and spatial memory. Grid cells exist in the medial entorhinal cortex of mammals, which is the area of the human brain frequently affected in early stages of Alzheimer’s disease, which deals with the loss of spatial memory (Moser et al., 2014). Finding the analogous region in the fish brain is important because it would provide an evolutionary link to the development of the mammalian cortex and serve as a somewhat simpler model in understanding the loss of spatial memory, with the hope one day of unlocking the mystery of Alzheimer’s disease.
Bibliography
Benhamou, Simon. “Place Navigation in Mammals: A Configuration-Based Model.” Animal Cognition 1 (1998): 55-63.
Broglio, Christina, et al. “Selective Involvement of the Goldfish Lateral Pallium in Spatial Memory.” Behavioral Brain Research 210 (2010): 191-201.
Brown, Culum. “Fish Intelligence, Sentience, and Ethics.” Animal Cognition 18.1 (1 Jan. 2015): 1-17.
Bshary, Redouan, Wolfgang Wickler, and Hans Fricke. “Fish Cognition: A Primate’s Eye View.” Animal Cognition 5 (2002): 1-13.
Burgess, Neil. “Spatial Memory: How Egocentric and Allocentric Combine.” Trends in Cognitive Sciences (2006). doi:10.1016/j.tics.2006.10.005.
Fish Net. Web. http://www.fishnet.org.
Hurtado-Parrado, Camilo. “Neuronal Mechanisms of Learning in Teleost Fish.” Universitas Psychologica 9.3 (2010): 663-678.
Ingle, David, and Dianne Shagian. “Solution of a Spatial Constancy Problem by Goldfish.” Physiological Psychology 1.2 (1973): 83-84.
Kandel, Eric R. In Search of Memory: The Emergence of a New Science of Mind. New York: W.W. Norton & Company, Inc., 2006.
Knierin, James J., and Derek A. Hamilton. “Framing Spatial Cognition: Neural Representations of Proximal and Distal Frames of Reference and Their Roles in Navigation.” Physiological Review 91 (2011): 1245-79.
López, Juan Carlos, et al. “Multiple Spatial Learning Strategies in Goldfish (Carassius auratus).” Animal Cognition 2.2 (1999): 109-120.
López, Juan Carlos, et al. “Reversal Learning Deficit in a Spatial Task But Not in a Cued One After Telencephalic Ablation in Goldfish.” Behavioral Brain Research 109 (2000): 91-98.
López, Juan Carlos, et al. “Dissociation of Place and Cue Learning by Telencephalic Ablation in Goldfish.” Behavioral Neuroscience 114.4 (2000): 687-699.
Moser, Edvard I., et al. “Grid Cells and Cortical Representation.” Nature 15 (2014): 466-481.
Nadel, Lynn. “The Hippocampus and Space Revisited.” Hippocampus 1.2 (1991): 221-239.
Odling-Smee, Lucy, and Victoria A. Braithwaite. “The Role of Learning in Fish Orientation.” Fish and Fisheries 4 (2003): 235-246.
Osborne, Darren. “Three-Second Fish Memory ‘Rubbish.’” News in Science, Australian Broadcasting Corporation. 14 Jan. 2010. Web. http://www.abc.net.au/science/articles/2010/01/14/2792407.htm.
Portavella, Manuel, and Juan P. Vargas. “Emotional and Spatial Learning in Goldfish is Dependent on Different Telencephalic Pallial Systems.” European Journal of Neuroscience 21 (2005): 2800-06.
Rodríguez, Fernando, et al. “Cognitive and Emotional Functions of the Teleost Fish Cerebellum.” Brain Research Bulletin 66.4-6 (2005): 365-370.
Rodríguez, Fernando, et al.. “Conservation of Spatial Memory Function in the Pallial Forebrain of Reptiles and Ray-Finned Fishes.” The Journal of Neuroscience 22.7 (2002): 2894-03.
Rodríguez, Fernando, et al. “Performance of Goldfish Trained in Allocentric and Egocentric Maze Procedures Suggests the Presence of a Cognitive Mapping System in Fishes.” Animal Learning and Behavior 4 (1994): 409-420.
Saito, Kotaro, and Shigeru Watanabe. “Experimental Analysis of Spatial Learning in Goldfish.” The Psychological Record 55 (2005): 647-662.
Salas, Cosme, et al. “Telencephalic Ablation in Goldfish Impairs Performance in a ‘Spatial Constancy’ Problem But Not in a Cued One.” Behavioral Brain Research 79 (1996): 193-200.
Salas, Cosme, et al. “Neuropsychology of Learning and Memory in Teleost Fish.” Zebrafish 3.2 (2006): 157-171.
Salas, Cosme, et al. “Spatial Learning and Memory Deficits After Telencephalic Ablation in Goldfish Trained in Place and Turn Maze Procedures.” Behavioral Neuroscience 110.5 (1996): 965-980.
Shubin, Neil. Your Inner Fish: A Journey into the 3.5-Billion-Year History of the Human Body. New York: Vintage Books, 2009.
Tolman, Edward C. “Cognitive Maps in Rats and Men.” Psychological Review 55.4 (1948): 189-208.
Tolman, E.C., B.F. Ritchie, and D. Kalish. “Studies in Spatial Learning: II. Place Learning Versus Response Learning.” Journal of Experimental Psychology 36 (1946): 221-229.
Tolman, E.C., B.F. Ritchie, and D. Kalish. “Studies in Spatial Learning: I. Orientation and the Shortcut.” Journal of Exploratory Psychology 1 (1946): 13-24.
Vargas, Juan Pedro, et al. “Encoding of Geometric and Featural Spatial Information by Goldfish (Carassius auratus).” Journal of Comparative Psychology 118.2 (2004): 206-216.